direkt in Ihrer Tasche








Zusammenfassung
Die Doppelfüßer (Diplopoda) sind eine Klasse der Gliederfüßer (Arthropoda) und werden bei den Tausendfüßern (Myriapoda) eingeordnet. Weltweit sind über 12.000 Arten dieser Tiere bekannt, damit stellen sie die größte Gruppe der Tausendfüßer dar. Es wird jedoch davon ausgegangen, dass etwa 15.000–80.000 Arten existieren. In Europa leben etwa 1500 Arten, in Deutschland sind es etwa 150. Tausendfüßer gibt es bereits seit rund 410 Millionen Jahren (Silur); sie gehörten zu den ersten Landbewohnern und stellen damit eine sehr urtümliche Tiergruppe dar. Arten der Gattung Arthropleura lebten vor 310 Millionen Jahren und erreichten eine Länge von mitunter zwei Metern. Damit waren es die größten bislang bekannten Landarthropoden, die jemals auf der Erde lebten. Wenn umgangssprachlich von Tausendfüßern die Rede ist, sind meistens Doppelfüßer, vor allem Vertreter der Ordnung Schnurfüßer (Julida) und ähnliche Ordnungen (Spirobolida, Spirostreptida), gemeint.
Species
Bumblebee Millipede
Anadenobolus monilicornis
American Giant Millipede
Narceus americanus
Black-and-gold Flat Millipede
Apheloria virginiensis
Florida Ivory Millipede
Chicobolus spinigerus
Paeromopus angusticeps
Paeromopus angusticeps
Rudiloria trimaculata
Rudiloria trimaculata
Portuguese Millipede
Ommatoiulus moreleti
Rusty Millipede
Trigoniulus corallinus
Yellow-legged Pleuroloma
Pleuroloma flavipes
Pseudopolydesmus serratus
Pseudopolydesmus serratus
Striped Millipede
Ommatoiulus sabulosus
Yellow-and-black Flat Millipede
Apheloria tigana
Narceus annularis
Narceus annularis
Pseudopolydesmus canadensis
Pseudopolydesmus canadensis
Desert Millipede
Orthoporus ornatus
Segmentierung und Grundbauplan
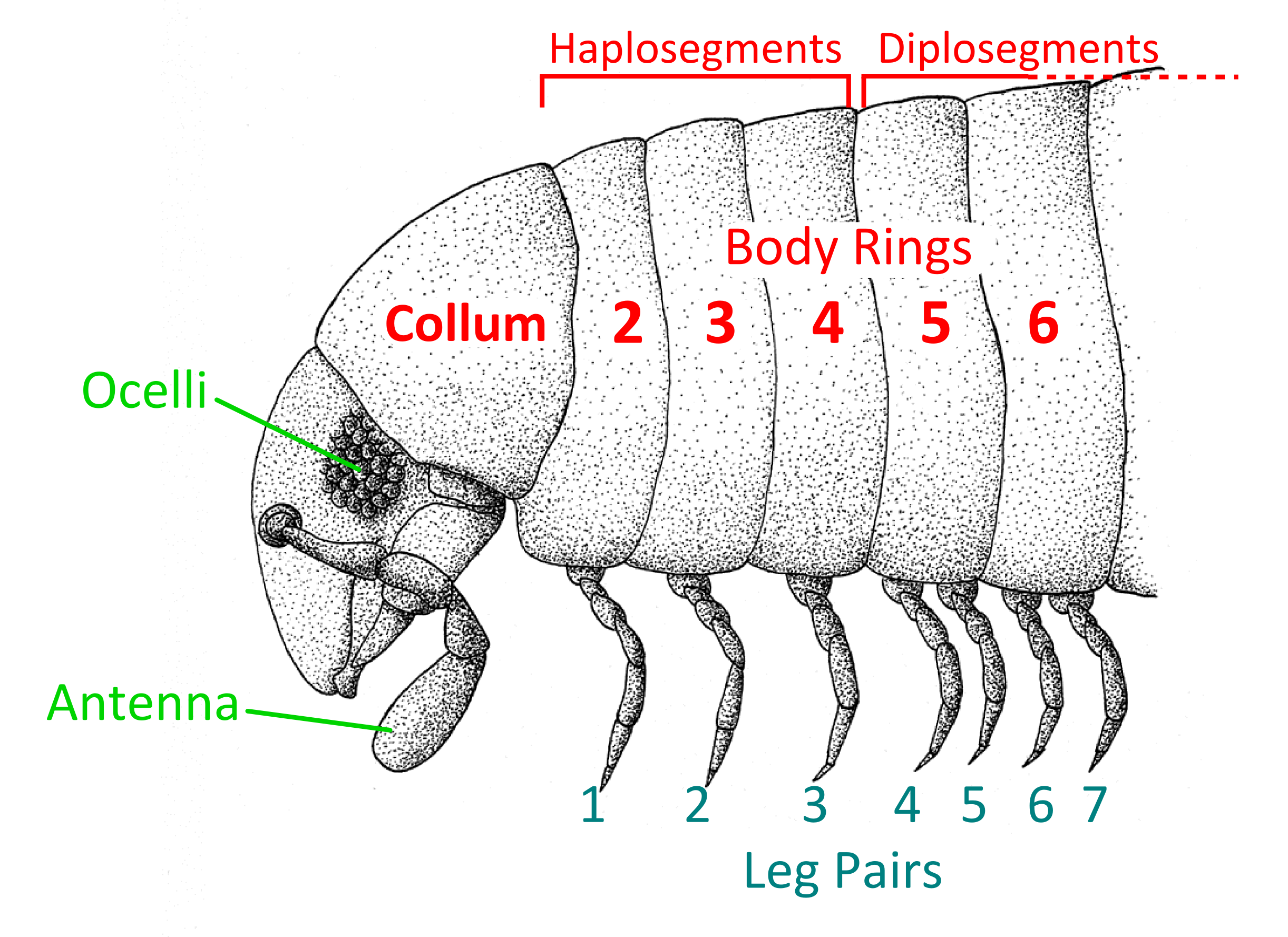
Wie alle Angehörigen der Myriapoden zeichnen sich die Doppelfüßer vor allem durch eine einheitliche Gliederung der Körpersegmente aus. Auf den Kopf folgen die Rumpfsegmente, von denen der Halsschild (Collum) das erste darstellt. Der Halsschild trägt keine Beine. Das auffälligste und namensgebende Merkmal der Gruppe ist eine Verschmelzung der Segmente (beginnend vom 5. Rumpfsegment, also dem 6. Segment des Körpers insgesamt) zu Doppelsegmenten, wodurch jedes dieser Diplosegmente zwei Beinpaare besitzt. Dadurch ist häufig auch von Körperringen die Rede, da es sich bei den Diplosegmenten nicht um Segmente im streng wissenschaftlichen Sinne handelt, sondern um je zwei verschmolzene Segmente. So besteht z. B. der Körper eines typischen Bandfüßers aus 20 Körperringen, nicht aber 20 echten Segmenten. Dennoch wird der Begriff Segment häufig auch für die einzelnen sichtbar getrennten Teile des Körpers verwendet (Kopf – Halsschild – Haplosegmente des Rumpfes – Doppelsegmente des Rumpfes). Der schmalere vordere Teil eines Doppelsegments ist das Prozonit, der hintere, etwas breitere und geriffelte, das Metazonit. Seitenflügel (Paranota, auch Paraterga genannt), Seitenkiele und ähnliche Strukturen liegen dabei meistens am Metazoniten. Die Verschmelzung der einzelnen Segmentteile erfolgt in den einzelnen Ordnungen unterschiedlich, was auch die unterschiedlichen Körperformen der Doppelfüßer bedingt (im Querschnitt kreisrund bis stark abgeplattet). Vor allem die Doppelfüßer werden oft mit der Bezeichnung "Tausendfüßer" gemeint, da sie von allen Gruppen der Myriapoden die meisten Beinpaare besitzen. Die minimale Anzahl an Beinpaaren bei adulten Tieren liegt bei 13, die maximale Anzahl der Beinpaare lag lange Zeit bei "nur" 375 Paaren (Illacme plenipes), also deutlich unter den 500 Beinpaaren, die für Tausend Beine notwendig wären. 2021 wurde jedoch mit Eumillipes persephone erstmals eine Art mit 1306 Beinen entdeckt. Die maximale Anzahl an Körperringen lag vor der Entdeckung von Eumillipes persephone bei 190, seit der Entdeckung liegt sie bei 330. Am Körperende befindet sich das Telson als letztes Segment des Körpers. Dabei handelt es sich beim Telson ebenso wenig wie beim Kopf um ein echtes Segment. Das Telson besteht zum einen aus einem beinlosen Präanalring, der aus einem Tergit auf der Oberseite besteht, das zu einem Schwänzchen (Epiproct) ausgezogen sein kann, und einem Sternit auf der Unterseite, der Analschuppe (Hypoproct, Subanalschuppe). Ein weiterer Bestandteil des Telsons ist ein Paar Paraprocta (im englischen anal valves genannt), die das Körperende bilden, an dem sich der Anus befindet. Vor dem Präanalring weisen die Jugendstadien eine mehr oder weniger große Anzahl beinloser Körperringe auf, die je nach Stadium und Art unterschiedlich ist. Die Atmung erfolgt über Tracheen. Die Tracheenöffnungen liegen knapp oberhalb der Beinbasen, beginnend am 3. Laufbeinpaar. Die Größe der Doppelfüßer reicht von wenigen Millimetern bis zu 30 cm.


Morphologie des Kopfes
Der stark gewölbte Kopf der Doppelfüßer besitzt sehr kleine Antennen, die nach unten gebogen werden und mit speziellen Sinnesorganen den Boden abtasten. Die Antennen der Doppelfüßer besitzen vier Sinneskegel (sense cones). Die Tiere besitzen an jeder Kopfseite Augenfelder, die aus Einzelaugen (Ocellen, auch Ommatidien genannt) bestehen, sowie ein Schläfenorgan (Tömösvárysches Organ) ebenfalls beiderseits des Kopfes. Bei den Penicillata finden sich außerdem Sinneshaare (Trichobothrien) am Kopf. Die kräftige Mandibel ist dreiteilig, der Mundraum wird hinten durch eine von der 1. Maxille gebildeten Unterlippe (Gnathochilarium) abgeschlossen. Die 2. Maxille bleibt rudimentär und bildet nur den Hinterrand der Unterlippe.

Exoskelett und Wehrdrüsen
Die ursprünglichsten Formen der Doppelfüßer (die Penicillata) besitzen maximal 17 Beinpaare und stellen sehr kleine, weichhäutige Formen dar. Bei den restlichen, abgeleiteten Ordnungen wird in die Chitinhaut des Exoskeletts Kalk zur Versteifung eingelagert. Dadurch sind sie stärker gepanzert, was ihnen Schutz und Stabilisierung beim Durchwühlen des Bodens, Schutz vor Austrocknung und Schutz vor Feinden bietet. An den Seiten des Körpers besitzen viele Doppelfüßer der Unterklasse Chilognatha in oder an der Naht zwischen Pro- und Metazonit Wehrdrüsen, aus denen giftige Abwehrsekrete abgegeben werden können. Diese Sekrete fallen oft durch einen penetranten Geruch auf, der sogar bei einem Waldspaziergang wahrgenommen werden kann, wenn sich Doppelfüßer dort durch Schritte im Laub gestört fühlen und das Sekret absondern. Die Wehrdrüsenöffnungen (Ozoporen) sind bei einigen Arten als Punkte an den Körperseiten sichtbar. Im Fall der Glomerida enthält das Wehrsekret u. a. die Chinazolin-Alkaloide Glomerin und Homoglomerin. Julidae bilden in ihren Wehrdrüsen Benzochinone, Spirobolida bilden Benzochinone und Hydrochinon und die Polydesmida freie Blausäure und Benzaldehyd. Viele Arten können diese Sekrete über mehrere Zentimeter verspritzen.
Geschlechtsmerkmale
Die Geschlechtsöffnung befindet sich bei allen Doppelfüßern hinter dem 2. Laufbein, wobei die Männchen zwei kompliziert gebaute Penes besitzen. Die weiblichen Tiere besitzen Vulven (Gonoporen). Die Männchen der meisten Arten haben außerdem zu Begattungsbeinen umgewandelte Laufbeine, die Gonopoden oder Kopulationsfüße, mit denen das Sperma vom Penis in die Vulva des Weibchens befördert werden kann. Diese sitzen meist am 7. Körperring, können sich bei den Pentazonia aber auch am Körperende befinden und dann Telopoden genannt werden. Für die Bestimmung vieler Arten ist die Form der Gonopoden wichtig, da die sonstigen äußeren Merkmale oftmals identisch sind. Die Spermien der Doppelfüßer besitzen kein Flagellum (Geißel), was neben den verschmolzenen Doppelsegmenten eine weitere Apomorphie der Klasse darstellt.
Merkmale der einzelnen Gruppen (Taxa) der Doppelfüßer
Die Diplopoda werden in zwei rezente Unterklassen unterteilt: Die Penicillata und die Chilognatha. Zu den Penicillata gehören einzig die Pinselfüßer, die Chilognatha beinhalten alle sonstigen Ordnungen. Folgende Merkmale dienen der Differenzierung: Die Chilognatha sind wiederum in zwei Infraklassen aufgeteilt: Die Pentazonia und die Helminthomorpha (wurmartigen Doppelfüßer). Diese lassen sich wie folgt morphologisch erkennen: Die Pentazonia sind in die Überordnungen Limacomorpha und Oniscomorpha unterteilt. Die Limacomorpha enthalten dabei nur die Urtausendfüßer (Glomeridesmida). Die Oniscomorpha („Asselähnlichen“, Kugler) enthalten die Saftkugler (Glomerida) und die Riesenkugler (Sphaerotheriidae). Für einen Vergleich der Ordnungen der Pentazonia siehe folgende Tabelle: Die Antennen der Saftkugler stehen zudem näher beisammen als die der Riesenkugler. Die Helminthomorpha werden dagegen in zwei Teilklassen unterteilt: Die Colobognatha (saugende Doppelfüßer) und die Eugnatha. Diese unterscheiden sich wie folgt: Für einen Vergleich der Ordnungen der Colobognatha siehe folgende Tabelle: Die Eugnatha sind wiederum in mehrere Überordnungen aufgeteilt, wie die Nematophora, die Juliformia und die Merocheta. Folgende Merkmale charakterisieren diese Taxa: Für einen Vergleich der Ordnungen der Nematophora siehe folgende Tabelle. Die in der Tabelle aufgelisteten Ordnungen weisen alle eine dorsale Rille auf und in jeder der Ordnungen besitzen viele Arten Augen. Für einen Vergleich der Ordnungen der Juliformia siehe folgende Tabelle: Die Merocheta enthalten nur eine Ordnung, die Polydesmida (Bandfüßer). - Penicillata: weiche Außenhaut (Cuticula) ohne Kalkeinlagerungen; keine Gonopoden oder Telopoden; keine Wehrdrüsen; in „Pinseln“ angeordnete Haare; Quernaht zwischen Antennen und Ocellen; Trichobothrien am Kopf; Tergite, Pleurite und Sternite durch Membranen verbunden; 11–13 Körperringe; 13–17 Beinpaare, keine Kopulation, große Taster am Gnathochilarium; Tömösvárysches Organ klein - Chilognatha: kalzifizierte Cuticula; ohne Trichobothrien; Coxae des ersten Beinpaars der Weibchen mit den Sterniten verbunden - Pentazonia: geteilte Sternite; Labrum mit einem einzigen medianen Zahn; Tergit des Telsons vergrößert und bedeckt das Analsegment; Kopfkapsel ohne mediane Quernaht; Tömösvárysches Organ vorhanden; Tergite, Pleurite und Sternite frei; Tracheenöffnungen am 2. und 3. Körperring; mediane Längsnaht der Tergite nicht vorhanden; zumindest das letzte Beinpaar der Männchen zu Telopoden umgewandelt; verzweigte Tracheen - Helminthomorpha: laterale Reihe der Ozoporen beginnt ab dem 5. Körperring; keine Tracheenöffnungen am 2. und 3. Körperring; keine verzweigten Tracheen; Tergite bestehen aus Prozonit und Metazonit, keine geteilten Sternite; ohne Telopoden am Körperende, dafür mit Gonopoden am vorderen Bereich des Körpers; - Colobognatha: Männchen mit 2 Paaren beinähnlicher Gonopoden (L9 zur Spermaübertragung + L10) mit einer variablen Anzahl an Podomeren am 7. und 8. Körperring, davor 8 Beinpaare, die Gonopoden entstehen im Verlauf mehrerer Häutungen sukzessive aus Laufbeinen; Gnathochilarium ohne Taster; Kopf schmaler als der restliche Körper; maximal ein oder zwei Paare an Ocellen; Tömösvárysches Organ nicht vorhanden; Subanalplatte nicht vorhanden - Eugnatha: Männchen mit einem Paar Gonopoden (L8) am 7. Körperring (L8 = anteriores Beinpaar), davor 7 Beinpaare, Gonopoden entstehen ontogenetisch aus knospenartigen Gebilden; Tergite und Pleurite verschmolzen; kugelförmige Wehrdrüsen; Gnathochilarium mit Tastern; Tracheenöffnungen oftmals verschließbar - Nematophora: Spinngriffel am Körperende; Tracheen die zu Spingriffeln führen verzweigt; Sternite frei - Juliformia: Sternite sind mit den Pleurotergiten verbunden, aber durch eine Naht abgegrenzt; Collum vergrößert und bedeckt posterioren Teil der Kopfkapsel; Spermium mit Pseudoperforatorium; Tömösvárysches Organ nicht vorhanden; produziert Wehrsekrete mit Benzoquinonen; L8 und L9 Gonopoden - Merocheta: 19–21 Körperringe; fingerförmiges Organ am 7. Antennomer; Sternite ohne Naht verbunden; keine Ocellen; keine Ozoporen am 6. Körperring; birnenförmiges Tömösvárysches Organ, L9 unmodifiziert
Fortpflanzung
Innerhalb der Doppelfüßer gibt es ursprüngliche Arten mit indirekter sowie weiter entwickelte Arten mit direkter Spermienübertragung. Die Männchen der Penicillata spinnen ein Fadenkonstrukt, auf dem sie einen Spermatropfen platzieren. Mit Hilfe von Signalfäden finden die Weibchen das Gespinst und nehmen das Sperma auf. Diese Form der Befruchtung entspricht weitestgehend der der Wenigfüßer. Bei allen anderen Doppelfüßern kommt es zu einer direkten Begattung der Weibchen durch die Männchen. Die Spermienübertragung erfolgt durch speziell ausgebildete Kopulationsorgane, die innerhalb der Gruppen auf unterschiedliche Weise aus Extremitäten entwickelt wurden. So haben die Vertreter der Pentazonia speziell umgebildete Endbeine (Telopoden), mit denen sie das Weibchen festhalten und begatten können. Die Männchen der Helminthomorpha besitzen mehrere vordere Laufbeine (im Bereich des 7. bis 11. Laufbeinpaares), die umgebildet wurden (Gonopoden). Auch Parthenogenese ist bei vielen Arten der Doppelfüßer nachgewiesen worden. Die Eier legt das Weibchen in Erdritzen ab oder platziert sie in Erdkämmerchen. Die Nematophora umgeben sie mit einem Gespinst und innerhalb der Colobgnatha findet man auch Brutpflege. Dabei rollen sich die Weibchen und (seltener) auch die Männchen um die Eier.
Entwicklung vom Ei zum Adulten
Die Eier der Doppelfüßer sind rundlich bis oval, meist hellgelblich bis weiß gefärbt und werden von einer festen, lederartigen Eihülle, dem Chorion, umgeben. Der Embryo benötigt mehrere Wochen zu seiner Entwicklung. Bei manchen Arten, z. B. Vertretern der Polydesmida, besitzt er einen Eizahn oder ein ganzes Feld davon, welche dabei helfen, das Chorion zu öffnen. Bei den meisten Arten geschieht dies aber lediglich durch einen erhöhten Innendruck, der die Eihülle zum Aufplatzen bringt. Daraus hervor quillt ein kommaförmiges, an eine Insektenpuppe erinnerndes Gebilde, das Pupoid. Es ist wiederum noch von einer dünnen Hautschicht umgeben. Nach Abstreifen der Hülle schlüpft ein mit drei Beinpaaren und einem viergliedrigen Antennenpaar ausgestattetes Jungtier, das Stadium I. Von den mitteleuropäischen Ordnungen läuft es nur bei den Pinselfüern und Saugfüßern bereits herum und ist zu selbstständiger Nahrungsaufnahme befähigt. Bei den Saftkuglern verbleibt es in der Eikapsel, die Jungtiere der Bandfüßer und Schnurfüßer verbleiben in ihren Nestern. Sie ernähren sich noch vom Dottervorrat. Das Stadium I der Schnurfüßer ist kommaförmig gekrümmt, rein weiß mit einem roten Ommatidium (Augenfleck) und fast unbeweglich. Erst Stadium II beginnt zögerlich herumzulaufen und kann somit, wenn auch eher selten, in Bodenfallenfängen nachgewiesen werden. Die Doppelfüßer entwickeln sich durch Häutungen, wobei mit jeder Häutung die Größe, die Zahl der Körperringe und der Beine zunimmt und sich immer mehr Strukturen ausprägen, z. B. die Geschlechtsmerkmale. Das Gesetz der Anamorphose wird hier am Beispiel der ringbildenden Juliformia erläutert. Das Gesetz besagt: Alle beinlosen Körperringe eines bestimmten Stadiums werden im folgenden Stadium zu beinpaartragenden Ringen. Sie tragen im vorausgehenden Stadium jedoch schon kleine Beinknospen. Gefolgt werden die beinlosen Körperringe von einer Zone mit frühen Ansätzen der zukünftigen beinlosen Körperringe. Meist ist diese Zone heller gefärbt und wenig sklerotisiert (verhärtet). Entsprechend dem normalen Bauplan der Schnurfüßer schließt sich daran die Sprossungszone an, von der aus die neuen Ringanlagen gebildet werden. Das Telson bildet das Körperende. Bei der Häutung in Stadium II werden die zwei im Stadium I beinlosen Körperringe zu beinpaartragenden, so dass das Tier jetzt 5 beinpaartragende und 4–5 beinlose Körperringe besitzt usw. Bei Schnurfüßern mit deutlichen Wehrdrüsenpunkten lässt sich die Entwicklung auch sehr gut an diesen verfolgen. Die erste funktionsfähige Wehrdrüse tritt im Stadium II am 2. beinpaartragenden Körperring (Körperring 6) auf, gefolgt von den schon besprochenen 4–5 beinlosen Körperringen, die nur mit den Anlagen von Wehrdrüsen ausgestattet sind. Erst bei der nächsten Häutung in Stadium III werden diese funktionsfähig und bei den meisten Arten durch Wehrdrüsenflecken (charakteristische Pigmentierung im Umfeld der Wehrdrüsen) gut sichtbar. Die zuletzt angelegten Wehrdrüsenflecken sind kleiner und heller gefärbt, die älteren größer und dunkler. Von der Anzahl der erkennbaren Serien kann man Rückschlüsse auf das Alter der Tiere ziehen, wobei allerdings beachtet werden muss, dass sich die Größen- und Farbunterschiede alter Wehrdrüsenserien stark angleichen und sich dann nicht mehr unterscheiden lassen. Aber auch das Augenfeld bietet, sofern vorhanden, die Möglichkeit, das stadiale Wachstum der Tiere zu verfolgen. Ausgehend von einem Ommatidium im Stadium II nimmt bei jeder Häutung die Zahl der Augenreihen kontinuierlich zu. Stadium III besitzt das alte Ommatidium plus eine Reihe mehr, bestehend aus 2 Ommatidien. Stadium IV hat 1+2+3 Ommatidien usw. Dies setzt sich solange fort, bis das Augenfeld durch das Halsschild begrenzt wird. Dies ist natürlich nur für die Arten der Fall, bei denen solch ein aus vielen Ommatidien bestehendes, dreieckiges Augenfeld vorhanden ist. Viele Arten besitzen auch nur ein geringe Anzahl an Ommatidien. Jungtiere der Doppelfüßer unterscheiden sich also meist von den Erwachsenen erstens in der Gesamtzahl der Körperringe bzw. Wehrdrüsenflecken und zweitens in der hohen Zahl (4–8) beinloser Ringe am Körperende. Außerdem besitzen sie viel weniger Ommatidien. Da es aber schwierig ist diese Merkmale mit einer Lupe am lebenden Tier zu erkennen oder die Ringe abzuzählen, kann es leicht zu Verwechslungen ähnlicher Arten kommen, wenn Jungtiere vorliegen.

Alter und Entwicklungsformen
Das Alter von Doppelfüßern kann stark variieren. So sind viele Samenfüßer in kälteren Gebieten nur einjährig und leben vom Schlupf bis zum Tod nur ein halbes Jahr lang, maximal knapp über ein Jahr. Neben den einjährigen Arten gibt es aber auch mehrjährige Arten, die den größeren Anteil der Arten ausmachen. Sehr viele Arten häuten sich Zeit ihres Lebens weiter, in der Regel findet nach dem Erreichen der Geschlechtsreife eine Häutung pro Jahr statt, die jeweils mit einem Körperringzuwachs und anschließender Bildung der Geschlechtsprodukte verbunden ist. Ein solcher Lebenszyklus findet sich zum Beispiel bei Schnurfüßern oder Saugfüßern und wird als Euanamorphose bezeichnet. Diese Arten können ein erstaunlich hohes Alter von bis zu 17 Jahren erreichen. Einige Arten der Schnurfüßer haben eine zusätzliche Strategie zur Lebensverlängerung entwickelt. Bereits geschlechtsreife Männchen können sich bei einer folgenden Häutung in ein präadultes (nicht erwachsenes) Individuum zurückverwandeln, bei einer weiteren Häutung wieder zu einem adulten Tier werden oder sich nochmals zu einem präadulten häuten. Weil es hier zu einem periodischen Wechsel zwischen geschlechtsreifen und sexuell inaktiven Stadien kommt, spricht man von Periodomorphose. Die Ursache für solche dazwischen geschalteten Stadien („Schaltmännchen“) ist bisher unbekannt. Auslöser scheinen veränderte Umweltbedingungen zu sein oder aber der Drang, bei zu hohen Individuendichten der Art im Habitat, sich einen neuen Lebensraum zu erschließen. Nicht geschlechtsreife Tiere eignen sich dafür besser als adulte, da sie ihre Energie nicht in das Fortpflanzungsgeschehen investieren müssen, sondern sich auf die Lebensraumerweiterung durch weite Wanderungen konzentrieren können. Es gibt aber auch Arten, die sich ebenfalls Zeit ihres Lebens häuten, dabei aber keinen Zuwachs an Körperringen bzw. Beinpaaren erhalten. Sie nehmen bei jeder Häutung nur in der Größe zu. Diese Entwicklung wird als Hemianamorphose bezeichnet und findet sich bei den Pinselfüßern und den Kuglern. Kugler können bis zu 11 Jahre alt werden. Bei den Bandfüßern und Samenfüßern ist das Erreichen der Geschlechtsreife das Endstadium der Entwicklung (Teloanamorphose). Sie haben unter den Doppelfüßern die kürzeste Lebenserwartung. Die Tiere sterben nach erfolgter Fortpflanzung (meist) im 2. oder 3. Lebensjahr ab.



Verbreitung
Doppelfüßer sind nahezu weltweit verbreitet. Sie kommen auf allen Kontinenten außer der Antarktis vor. Weitere Verbreitungslücken liegen beispielsweise in einem Großteil der Sahara und der Arabischen Halbinsel, in einigen Wüsten Zentralasiens und in den arktischen Gebieten der Nordhalbkugel. In Europa finden sie sich nicht nördlich des 70. Breitengrades, in Asien nicht nördlich des 67. Breitengrades, in Alaska nicht nördlich des 66. Breitengrades und im Osten Nordamerikas nicht nördlich des 55. Breitengrades. Dabei können Ausnahmen durch Verschleppungen häufig auftreten. Auch auf den meisten größeren Inseln der Welt leben Doppelfüßer. Bei der Verbreitung der Doppelfüßer zeigen sich zwei Verbreitungsschwerpunkte: Zum einen die Tropen und hier insbesondere tropische Regenwälder und zum anderen Gebirge bzw. Gebirgswälder generell – so ist eine beeindruckende Artenzahl aus den Alpen, Pyrenäen oder Appalachen bekannt, die in der subtropischen oder gemäßigten Zone liegen. Auch aus gebirgigen Gegenden in den Tropen sind viele Arten bekannt, z. B. den Anden oder dem Himalaya. Da viele Ordnungen ein hohes Feuchtigkeitsbedürfnis haben (also hygrophil sind), bevorzugen sie feuchte Lebensräume. Diese sind beispielsweise im humiden Ostseitenklima häufig zu finden, weshalb es in Ostasien, dem Osten Nordamerikas oder Südosteuropa eine hohe Zahl an Taxa und Arten gibt. Aber auch in trockenem Westseitenklima tritt regional eine hohe Artenzahl auf, beispielsweise im Mittelmeerraum oder Kalifornien. Viele Arten weichen in heißtrockenen Klimaten auch in Höhlen aus, manche Arten haben sich aber auch an Trockenheit angepasst. In Europa sind circa 1500 Arten von Doppelfüßern verbreitet, darunter 600 in der Ordnung Julida und über 500 in der Ordnung Chordeumatida.
Lebensweise und Ernährung
Die meisten Doppelfüßer leben im Boden (euedaphisch), in der Humusschicht (hemiedaphisch), in der Laubstreu/Streuschicht (epedaphisch), in zerfallendem Holz oder in Höhlen. Einige Arten klettern auch an Bäumen hoch oder leben vollständig auf Bäumen, wie die Pinselfüßer. In den meisten Fällen leben sie jedoch in Bodennähe und bilden einen Teil des Edaphons. Die meisten Doppelfüßer sind nachtaktiv. Dies dient vor allem als Schutz vor Austrocknung, die auch mit steigenden Temperaturen schneller eintritt. Tagsüber finden sich viele Arten unter Totholz, Steinen, in alten Baumstubben, im Bodeninneren oder an anderen geschützten Plätzen. Die Nahrung besteht meistens aus totem Pflanzenmaterial (Laubstreu, abgestorbene Pflanzenteile etc.), Totholz (vor allem morschem Holz), Pilzen (v. a. Pilzhyphen, wie in weißfaulem Holz, seltener Fruchtkörper), seltener auch aus lebenden Pflanzenteilen (Gras, Gemüse etc.), Bakterienrasen, Aas oder in seltenen Fällen lebenden Kleintieren. Als Destruenten sind sie meistens nicht sehr wählerisch. Jedoch können einige Ordnungen aufgrund der saugenden Mundwerkzeuge eher Pilze und Bakterienrasen fressen, während Ordnungen mit kauend-beißenden Mundwerkzeugen auch Totholz und abgestorbenes Laub fressen können. In den meisten Fällen gelten Doppelfüßer als Nützlinge, die organische Abfälle beseitigen, nur in wenigen Fällen sind sie als Schädlinge gefürchtet, die bei Nahrungsmangel z. B. Gemüsekulturen schädigen können.
Systematik der Doppelfüßer
Die Doppelfüßer bilden gemeinsam mit den Wenigfüßern (Pauropoda) die Dignatha aufgrund der Verschmelzung der basalen Glieder der 1. Maxille zu einer Unterlippe (Gnathochilarium), des Verlusts der 2. Maxille beziehungsweise der rudimentären Anlage derselben in der Embryonalentwicklung, der Genitalöffnungen im zweiten Segment, der Tracheenöffnungen nahe der Beine sowie eines Jungtiers mit nur drei Beinpaaren. Bei den Wenigfüßern ist die Antenne jedoch verzweigt, es finden sich keine Diplosegmente und das Collum zeigt Rudimente von Beinen. Mit den Zwergfüßern (Symphyla) bilden die Dignatha das Taxon Progoneata aufgrund der Darm- und Fettkörperbildung innerhalb des Dotters sowie dem Aufbau der Mechanorezeptoren (Trichobothrien). Dieser Gruppe werden gemeinhin die Hundertfüßer (Chilopoda) als Schwestergruppe gegenübergestellt. Intern werden die Doppelfüßer wie im Text bereits erwähnt in die ursprünglichen Penicillata sowie die abgeleiteten Chilognatha aufgeteilt. Innerhalb der Chilognatha gibt es eine weitere Aufspaltung in verschiedene Taxa. Das folgende Kladogramm gibt eine vereinfachte Übersicht über die Ordnungen der Doppelfüßer, einer klassischen Einordnung folgend: In einer Studie von 2007 wurden zum ersten Mal morphologische und molekulare Daten kombiniert verwendet, um die Phylogenie der Doppelfüßer korrekter darzustellen. Das folgende Kladogramm zeigt die modernere Einordnung der Doppelfüßer-Ordnungen. Die Stemmiulida nehmen dabei eine basalere Stellung innerhalb der Helminthomorpha ein und die Polydesmida zeigen eine nähere Verwandtschaft zu den Colobognatha als zu den übrigen Eugnatha.

Arten im deutschsprachigen Gebiet nördlich der Alpen
Da es zahlreiche südeuropäische Arten gibt, die z. B. im Kanton Wallis, Kanton Tessin oder in Kärnten vorkommen, aber sonst nicht weiter nördlich zu finden sind, wurde darauf verzichtet, alle Arten Deutschlands, Österreichs und der Schweiz aufzulisten. In dieser Liste vorhanden sind die Arten aus dem deutschsprachigen Teil der Schweiz exklusive Arten, die nur im Wallis oder Tessin vorkommen, alle Arten Deutschlands sowie die Arten Österreichs mit Ausnahme von Arten, die nur im Kärnten oder südlichen Teil der Steiermark oder des Burgenlandes vorkommen. Im Folgenden sind alle Doppelfüßer aus dem deutschsprachigen Gebiet nördlich der Alpen aufgelistet: - Penicillata - Pinselfüßer – Polyxenida - Polyxenidae - Kleiner Pinselfüßer (Polyxenus lagurus) - Chilognatha - Pentazonia - Saftkugler – Glomerida - Glomeridae - Geoglomeris subterranea - Vierbänder-Saftkugler (Glomeris connexa) - Schweizer Saftkugler (Glomeris helvetica) - Östlicher Sechsstreifen-Saftkugler (Glomeris hexasticha) - Westlicher Sechsstreifen-Saftkugler (Glomeris intermedia) - Bunter Saftkugler (Glomeris klugii) - Gerandeter Saftkugler (Glomeris marginata) - Geschmückter Saftkugler (Glomeris ornata) - Zweireihen-Saftkugler (Glomeris pustulata) - Vierreihen-Saftkugler (Glomeris tetrasticha) - Alpen-Saftkugler (Glomeris transalpina) - Trachysphaera costata - Trachysphaera gibbula - Trachysphaera schmidti - Glomeridellidae - Glomeridella minima - Helminthomorpha - Saugfüßer – Polyzoniida - Polyzoniidae - Polyzonium germanicum - Samenfüßer – Chordeumatida - Anthroleucosomatidae - Anamastigona pulchella (nur D) - Attemsiidae - Dendromonomeron oribates - Dimastosternum franzi (nur in Tirol) - Dimastosternum holdhausi (nur in den österreichischen Alpen) - Polyphematia moniliformis (nur A) - Syngonopodium aceris (nur A) - Syngonopodium cornutum (nur A) - Brachychaeteumatidae - Brachychaeteuma bagnalli - Brachychaeteuma bradeae - Brachychaeteuma melanops (nur D) - Chamaesoma broelemanni (nur im Grenzgebiet D-FR-LUX) - Craspedosomatidae - Atractosoma meridionale - Bergamosoma canestrinii - Bomogona helvetica (nur CH) - Bunter Höckersamenfüßer (Craspedosoma rawlinsii) - Craspedosoma taurinorum - Dactylophorosoma nivisatelles - Helvetiosoma helveticum - Julogona tirolensis - Listrocheiritium bohemicum (nur A) - Listrocheiritium cervinum - Listrocheiritium noricum - Listrocheiritium nubium (nur A) - Listrocheiritium septentrionale (nur A) - Listrocheiritium styricum (nur A) - Listrocheiritium sussurinum (nur A) - Megalosoma canestrinii - Nanogona polydesmoides - Ochogona brentana - Ochogona caroli - Ochogona elaphron (nur A) - Ochogona holdhausi (nur A) - Ochogona hanfi (nur A) - Ochogona pusilla (nur A) - Ochogona regalis - Ochogona triaina - Oroposoma granitivagum (nur CH) - Oroposoma ticinense (nur CH) - Pterygophorosoma alticolum - Pyrgocyphosoma titianum (Endemit im Schwarzwald) - Rhymogona montivaga - Rhymogona serrata - Rhymogona verhoeffi - Rhymogona wehrana - Stenalpium brentanum - Chordeumatidae - Chordeuma sylvestre - Melogona broelemanni - Melogona gallica - Melogona scutellaris (nur CH) - Melogona transsilvanica - Melogona voigti - Mycogona germanica - Orthochordeumella fulva - Orthochordeumella pallida - Haaseidae - Haasea cyanopida (nur A) - Haasea filicis (nur A) - Haasea flavescens - Haasea fonticulorum - Haasea germanica - Haasea gruberi (nur A) - Haasea inflata (nur A) - Xiphogona oribates - Xylophageuma vomrathi - Xylophageuma zschokkei - Hungarosomatidae - Hungarosoma bokori (nur A) - Mastigophorophyllidae - Haploporatia cervina (nur A) - Haploporatia eremita - Haploporatia similis (nur A) - Mastigona bosniensis - Mastigona mutabilis - Mastigophorophyllon saxonicum - Neoatractosomatidae - Pseudocraspedosoma grypischium - Trimerophorella rhaetica - Trachygonidae - Halleinosoma noricum - Trachygona capito (nur A) - Verhoeffiidae - Haplogona oculodistincta - Schnurfüßer – Julida - Schnurfüßer (Julidae) - Allajulus dicentrus (nur A) - Allajulus groedensis - Allajulus molybdinus (nur A) - Allajulus nitidus - Alpityphlus seewaldi (nur A) - Brachyiulus bagnalli (nur A) - Brachyiulus lusitans - Brachyiulus pusillus - Castaneoiulus salicivorus - Cylindroiulus arborum - Cylindroiulus boleti - Cylindroiulus britannicus - Gemeiner Feldschnurfüßer (Cylindroiulus caeruleocinctus) - Cylindroiulus fulviceps - Cylindroiulus italicus - Cylindroiulus latestriatus - Cylindroiulus luridus - Cylindroiulus meinerti - Cylindroiulus parisiorum - Gepunkteter Schnurfüßer (Cylindroiulus punctatus) - Cylindroiulus truncorum - Cylindroiulus verhoeffi - Cylindroiulus vulnerarius - Cylindroiulus zinalensis - Enantiulus nanus - Hypsoiulus alpivagus - Julus montivagus - Gemeiner Dunkler Schnurfüßer (Julus scandinavius) - Julus scanicus - Julus terrestris - Kryphioiulus occultus - Leptoiulus abietum (nur A) - Leptoiulus alemannicus - Leptoiulus belgicus - Leptoiulus bertkaui - Leptoiulus broelemanni - Leptoiulus cibdellus - Leptoiulus helveticus (nur CH) - Leptoiulus kervillei - Leptoiulus montivagus - Leptoiulus noricus - Leptoiulus proximus - Leptoiulus riparius - Leptoiulus saltuvagus - Leptoiulus simplex - Leptoiulus trilobatus - Leptoiulus vagabundus (nur A) - Megaphyllum projectum - Megaphyllum silvaticum - Megaphyllum sjaelandicum - Megaphyllum unilineatum - Ommatoiulus rutilans - Sandschnurfüßer (Ommatoiulus sabulosus) - Ophyiulus aspidiorum (nur A) - Ophyiulus germanicus - Ophyiulus nigrofuscus - Ophyiulus pilosus - Pachypodoiulus eurypus - Pteridoiulus aspidiorum - Rossiulus vilnensis - Styrioiulus pelidnus - Schwarzer Schnurfüßer (Tachypodoiulus niger) - Unciger foetidus - Unciger transsilvanicus - Xestoiulus imbecillus (nur A) - Xestoiulus laeticollis - Fadenfüßer (Blaniulidae) - Archiboreoiulus pallidus - Gefleckter Doppelfüßer (Blaniulus guttulatus) - Boreoiulus tenuis - Choneiulus palmatus - Nopoiulus kochii - Proteroiulus fuscus - Nemasomatidae - Nemasoma varicorne - Bandfüßer – Polydesmida - Polydesmidae - Gemeiner Kleiner Bandfüßer (Brachydesmus superus) - Großer Westlicher Bandfüßer (Polydesmus angustus) - Großer Östlicher Bandfüßer (Polydesmus complanatus) - Polydesmus coriaceus - Sommer-Bandfüßer (Polydesmus denticulatus) - Polydesmus edentulus - Höckriger Bandfüßer (Polydesmus inconstans) - Polydesmus monticola - Polydesmus susatensis – eingeschleppt - Polydesmus xanthocrepis (nur A) - Propolydesmus germanicus - Schweizer Bandfüßer (Propolydesmus helveticus) - Rundflügel-Bandfüßer (Propolydesmus testaceus) - Macrosternodesmidae - Macrosternodesmus palicola - Ophiodesmus albonanus - Pyrgodesmidae - Poratia digitata – eingeschleppt - Paradoxosomatidae - Gewächshaus-Tausendfüßer (Oxidus gracilis) – eingeschleppt - Stosatea italica – eingeschleppt - Strongylosoma stigmatosum



_(20746265600).jpg)



.jpg)







.JPG)

_02.jpg)
_millipede_(3680001399).jpg)
