directamente en tu bolsillo








Resumen
Drosophila melanogaster (en griego significa literalmente «amante del rocío de vientre negro»), también llamada mosca del vinagre o mosca de la fruta, es una especie de díptero braquícero de la familia Drosophilidae. Recibe su nombre debido a que se alimenta de frutas en proceso de fermentación tales como manzanas, bananas, uvas, etc. Es una especie utilizada frecuentemente en experimentación genética, dado que posee un reducido número de cromosomas (4 pares), breve ciclo de vida (15-21 días) y aproximadamente el 61 % de los genes de enfermedades humanas que se conocen tienen una contrapartida identificable en el genoma de las moscas de la fruta, y el 50 % de las secuencias proteínicas de la mosca tiene análogos en los mamíferos. Para propósitos de investigación, fácilmente pueden reemplazar a los humanos. Se reproducen rápidamente, de modo que se pueden estudiar muchas generaciones en un corto espacio de tiempo, y ya se conoce el mapa completo de su genoma. Fue adoptada como animal de experimentación genética por Thomas Morgan a principios del siglo XX. Sus 165 Mb de genoma (1 Mb = 1 millón de pares de bases) fueron publicados en marzo de 2000 gracias al consorcio público y la compañía Celera Genomics. Alberga alrededor de 13.600 genes.
Contenido
Desarrollo
De una célula derivan células hijas que generan una posible asimetría. Presenta una asimetría inicial en la distribución de sus componentes citoplasmáticos que da lugar a sus diferencias de desarrollo. En la ovogénesis se generan células foliculares, células nodrizas y el ovocito. La mosca de la fruta, a 29 °C, alcanza a vivir 30 días; y el desarrollo de huevo a adulto demanda 7 días. El El primordio desarrolla diferencias en los ejes: anteroposterior y dorsoventral. Una sucesión de acontecimientos derivados de la asimetría inicial del cigoto se traduce en el control de la expresión génica de forma que las regiones diferentes del huevo adquieren distintas propiedades. Esto puede ocurrir por la diferente localización de los factores de transcripción y traducción en el huevo o por el control diferencial de las actividades de estos factores. Después sigue otra etapa en la que se determinan las identidades de las partes del embrión: se definen regiones de las que derivan partes concretas del cuerpo. Los genes que regulan el proceso codifican reguladores de la transcripción y actúan unos sobre otros de forma jerárquica. También actúan sobre otros genes que son los que verdaderamente se encargan del establecimiento de este patrón (actúan en cascada). También hay que tener en cuenta las interacciones célula-célula ya que definen las fronteras entre los grupos celulares.

Estructura de un segmento
Hay 3 grupos de genes en función de sus efectos sobre la estructura de un segmento: - Genes maternos: expresados por la madre en la ovogénesis. Actúan durante o después de la maduración del ovocito. Un ejemplo es el gen bicoid. - Genes de segmentación: se expresan tras la fertilización. Se encargan del número y polaridad de los segmentos (hay 3 grupos que actúan secuencialmente para definir las partes del embrión). - Genes homeóticos: controlan la identidad de los segmentos (no el número, ni polaridad o tamaño).

Etapas del desarrollo
La siguiente etapa del desarrollo depende de los genes que se expresan en la mosca madre. Estos genes se expresan antes de la fertilización. Pueden dividirse en: Existen cuatro grupos de genes que intervienen en el desarrollo de las diferentes partes del embrión. Cada grupo se organiza en una vía diferente que presenta un orden concreto de actuación. Cada vía se inicia con hechos que tiene lugar fuera del huevo, lo que tiene como resultado la localización de una señal dentro de este. Estas señales (son proteínas que reciben el nombre de morfógenos) se distribuyen de forma asimétrica para cumplir funciones diferentes. Del eje antero-posterior se encargan 3 sistemas y del dorso-ventral se encarga uno: Todos los componentes de los cuatro sistemas son maternos por lo que los sistemas que establecen el patrón inicial dependen de sucesos anteriores a la fertilización. - Genes somáticos maternos: se expresan en células somáticas = células foliculares. - Genes de línea germinal materna: pueden actuar tanto en células nodriza como en el ovocito. - Sistema Anterior: responsable del desarrollo de cabeza y tórax. Se requieren productos de la línea germinal materna para situar al producto del gen bicoid en el extremo anterior del huevo. - Sistema Posterior: responsable de los segmentos del abdomen. Muchos productos intervienen en la localización del producto del gen nanos, que inhibe la expresión de hunchback en el abdomen. - Sistema Terminal: desarrollo de estructuras de los extremos no segmentados del huevo. Depende de los genes somáticos maternos (activan el receptor codificado por torso). - Sistema Dorso-ventral: se inicia por una señal desde una célula folicular de la cara ventral del huevo y se transmite a través del receptor codificado por el gen Toll. Esto produce la generación de un gradiente de activación del factor de transcripción producido por el gen Dorsal.
Desarrollo dorso-ventral
Existe una compleja interrelación entre oocito y células foliculares (genes del oocito son necesarios para el desarrollo de células foliculares y señales de estas, transmitidas al oocito, provocan el desarrollo de estructuras ventrales). Otra vía se encarga del desarrollo dorsal durante el crecimiento del huevo. Los sistemas funcionan por la activación de una interacción ligando-receptor que desencadena una vía de transducción. El proceso depende, en su inicio, del gen Gurken (que actúa también en diferenciación antero-posterior). El mRNA de Gurken se sitúa en la cara posterior del oocito haciendo que las células foliculares adyacentes se diferencien en células posteriores. Estas células devuelven una señal que desencadena la producción de una red de microtúbulos que es necesaria para la polaridad. La polaridad dorsoventral se establece cuando gurken llega a la cara dorsal del oocito (depende de la expresión de varios genes más). El producto de Gurken actúa como ligando interaccionando con el receptor (producto del gen Torpedo) de una célula folicular. La activación de este receptor desencadena una vía de señalización cuyo efecto final es el impedimento a que se desarrolle la cara ventral en la dorsal (se produce un cambio en las propiedades de las células foliculares de esta cara). El desarrollo de estructuras ventrales requiere genes maternos que establecen el eje dorso-ventral. El sistema dorsal es necesario para el desarrollo de estructuras ventrales (como mesodermo y neuroectodermo). Mutaciones en él, impiden el desarrollo ventral. La vía del desarrollo ventral, también se inicia en las células foliculares y finaliza en el oocito. En las células foliculares se producen una serie de señales que acaban generando un ligando par el receptor (producto del gen Toll = primer componente de la vía, que actúa dentro del oocito). Toll es el gen crucial en el transporte de la señal al interior del oocito. El resto de componentes del grupo dorsal codifican productos que o regulan o son necesarios para la acción de Toll. Toll es una proteína transmembrana (homóloga al receptor de la interleuquina 1). La unión de su ligando al receptor Toll, activa la vía que determina el desarrollo ventral. La distribución del producto de este gen es muy variable, pero solo induce la formación de estructuras ventrales en lugares adecuados (parece que solo se expresa producto activo en ciertas regiones). Tras la unión del ligando, el receptor Toll se activa en la cara ventral del embrión. Esta activación desencadena en una serie de procesos en los que intervienen los productos de otros genes y que termina en la fosforilación del producto del gen cactus que es el regulador final del factor de transcripción del gen Dorsal. En el citoplasma hay un complejo cactus-dorsal inactivo pero que al fosforilarse cactus libera a la proteína dorsal, que entra en el núcleo. La activación de toll lleva a la activación de dorsal. Se establece un gradiente de proteína dorsal en el núcleo que va del lado dorsal al ventral en el embrión. En la cara ventral, la proteína dorsal se libera hacia el núcleo pero en la dorsal, permanece en el citoplasma. La proteína dorsal activa a los genes Twist y Snail (necesarios para el desarrollo de estructuras ventrales) e inhibe a los genes Decapentaplegic y Zerknullt (necesarios para el desarrollo de estructuras dorsales). La interacción inicial entre gurken y torpedo lleva a la represión de la actividad de spatzle en la cara dorsal del embrión (ligando de toll). La proteína dorsal, situada en el núcleo, inhibe la expresión de dpp. De este modo, las estructuras ventrales se forman según un gradiente nuclear de la proteína dorsal y las estructuras dorsales según un gradiente de la proteína dpp. - En el eje dorso-ventral hay tres bandas bastante próximas que definen las regiones en las que se forman mesodermo, neuroectodermo y ectodermo dorsal (ordenadas de ventral a dorsal).
Historia de uso en análisis genético
D. melanogaster fue uno de los primeros organismos utilizados para el análisis genético, y en la actualidad es uno de los organismos eucarióticos más ampliamente utilizados y genéticamente más conocidos. Thomas Hunt Morgan comenzó a usar esta especie en la Universidad de Columbia en 1910 en un laboratorio conocido como "la sala de moscas". Él y sus colaboradores (incluso los famosos genetistas A.H. Sturtevant, Calvin Bridges, y H. J. Muller), comenzaron experimentos utilizando botellas de leche para criar moscas y lupas para observarlas. Las lupas fueron reemplazadas más tarde por microscopios de disección. Gracias a estas moscas pequeñas e inofensivas, Morgan y sus colaboradores dilucidaron muchos principios básicos de herencia, incluso la herencia ligada al sexo, epistasis, alelos múltiples y mapeo de genes. Dentro del análisis genético, se realizaron estudios de pérdida de función, que implicaban el silenciamiento de diversos genes para observar cual era la función que llevaban a cabo. De esta forma, el genoma de la mosca de la fruta fue alterado, en primer lugar, mediante el uso de mutágenos que ocasionaban cambios en su secuencia de ADN sin un estricto control experimental, como elementos genéticos transponibles, rayos X o químicos mutagénicos. Más adelante, se procedió al empleo de ARN interferente (RNAi) que, junto al sistema GAL4-UAS, permite la edición en un tejido determinado específicamente. En último lugar, y más recientemente, su genoma es estudiado mediante el uso del sistema CRISPR junto con la nucleasa Cas9, permitiendo una edición más sencilla, efectiva y asequible. Gracias a este descubrimiento, se ha procedido al desarrollo de nuevas nucleasas con funciones complementarias a Cas9 que permitan profundizar en la edición y estudio de este organismo mediante este sistema, como la nucleasa Cas12a (anteriormente conocida como Cpf1), y la nucleasa CasΦ.
La genética del comportamiento y la neurociencia
Seymour Benzer y otros han utilizado mutaciones que afectan el comportamiento de estas moscas para aislar genes implicados en la visión, el olfato, la audición, el aprendizaje, la memoria, el cortejo, el dolor, y otros procesos. Tras el trabajo pionero de Alfred Henry Sturtevant, Benzer y colegas utilizaron ginandromorfos (mosaicos sexuales) para desarrollar la nueva técnica de mapeo del destino. Esta técnica permitió asignar una característica particular a una ubicación anatómica específica. Por ejemplo, esta técnica demostró que el comportamiento de cortejo sexual masculino está controlado por el cerebro. El mapeo del destino del ginandromorfos también proporcionó la primera indicación de la existencia de feromonas en esta especie. Los machos distinguen entre machos y hembras conespecíficos y dirigen el cortejo hacia las hembras, gracias a una feromona sexual específica que las hembras producen principalmente en sus tergitos (placas dorsales endurecidas del exterior del abdomen).
Genoma
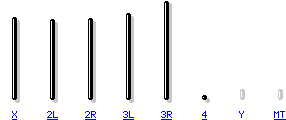
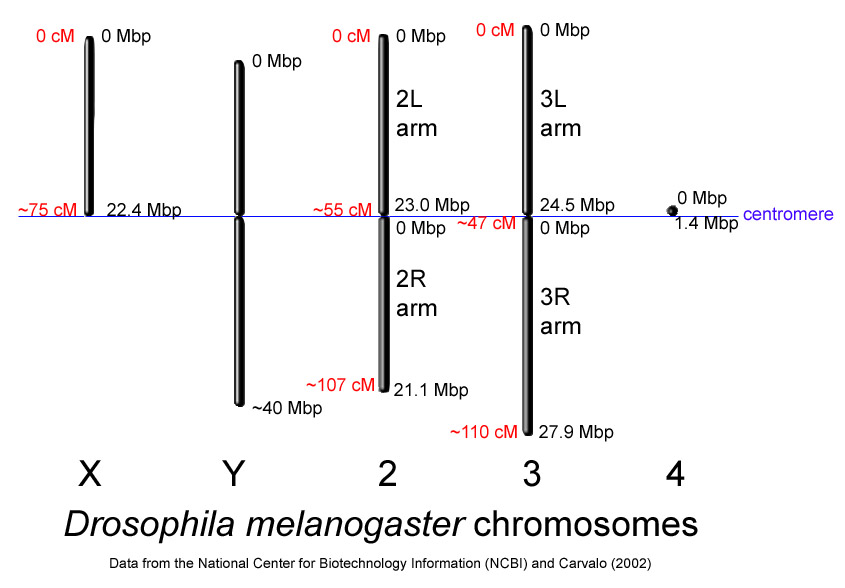
El genoma de D. melanogaster (secuenciado en 2000, y verificado en la base de datos FlyBase ) contiene cuatro pares de cromosomas: un par X/Y, y tres autosomas señalados como 2, 3, 4. El cuarto cromosoma es tan pequeño que a veces se ignora, salvo el importante gen sin ojos. El genoma secuenciado de D. melanogaster de 139,5 millones de pares de bases contiene aproximadamente 15.016 genes. Más del 60% de su genoma es funcional al codificar ADN no codificador de proteínas involucrados en el control de la expresión génica. La determinación de sexo en Drosophila se produce por la relación de cromosomas X a autosomas, no debido a la presencia de un cromosoma Y como ocurre en la determinación de sexo en humanos. Aunque el cromosoma Y es enteramente heterocromática, contiene al menos 16 genes, muchos de los cuales cumplen funciones relativas al sexo macho.
Similitud con humanos
Cerca del 75% de genes humanos vinculados con enfermedades, tienen su homólogo en el genoma de la mosca de la fruta, y el 50% de las secuencias de proteínas de la mosca tiene su homólogo en mamíferos. Existe una base de datos en línea, llamada Homophila está disponible para estudios de enfermedades genéticas humanas homólogas en moscas y viceversa. Drosophila sigue siendo usado extensamente como modelo genético para diversas enfermedades humanas incluyendo a desórdenes neurodegenerativos Parkinson, Huntington, ataxia espinocerebelosa y Alzheimer. Esta mosca también se usa en estudios de mecanismos del envejecimiento y estrés oxidativo, sistema inmunitario, diabetes, cáncer, abuso de drogas.
Enlaces externos
- Artículo sobre la investigación de la D. Melanogaster en la Estación Espacial Internacional - Berkeley Drosophila Genome Project - The Drosophila Genomics Resource Center - Mosca de la fruta en la Universidad de Kentucky - Subdirección de Moscas de la Fruta y Proyectos Fitosanitarios - Perú - Mediciones del cortejo en Drosophila melanogaster - Mantenimiento de Drosophila en lab: procedimientods generales - Transcripción In Situ de hibridación de embriones Whole-Mount para análisis fenotípico de RNAi-Treated Drosophila - Injection of dsRNA into Drosophila Embryos for RNA Interference (RNAi) - Genoma de Drosophila melanogaster, vía Ensembl - Genoma de Drosophila melanogaster (versión BDGP Release 6 + ISO1 MT/dm6), vía UCSC Genome Browser