directamente en tu bolsillo








Resumen
En la clasificación científica de los seres vivos, los animales (Animalia) o metazoos (Metazoa) constituyen un reino que reúne un amplio grupo de organismos que son eucariotas, heterótrofos, pluricelulares y tisulares (excepto los poríferos). Se caracterizan por su amplia capacidad de movimiento, por no tener cloroplasto (aunque hay excepciones, como en el caso de Elysia chlorotica) ni pared celular, y por su desarrollo embrionario; que atraviesa una fase de blástula y determina un plan corporal fijo (aunque muchas especies pueden sufrir una metamorfosis posterior como los artrópodos). Los animales forman un grupo natural estrechamente emparentado con los hongos (reino Fungi). Animalia es uno de los cinco reinos del dominio Eukaryota, y a él pertenece el ser humano. La parte de la biología que estudia los animales es la zoología. Los filos animales más conocidos aparecen en el registro fósil durante la denominada explosión cámbrica, sucedida en los mares hace unos 542 a 530 millones de años. Los animales se dividen en varios subgrupos, algunos de los cuales son vertebrados: (aves, mamíferos, anfibios, reptiles, peces) e invertebrados: artrópodos (insectos, arácnidos, miriápodos, crustáceos), anélidos (lombrices, sanguijuelas), moluscos (bivalvos, gasterópodos, cefalópodos), poríferos (esponjas), cnidarios (medusas, pólipos, corales), equinodermos (estrellas de mar), nematodos (gusanos cilíndricos), platelmintos (gusanos planos), etc.
Contenido
Species
Southern Black Widow
Latrodectus mactans
Western Black Widow
Latrodectus hesperus
Bumblebee Millipede
Anadenobolus monilicornis
California Ebony Tarantula
Aphonopelma eutylenum
American Giant Millipede
Narceus americanus
Florida Ivory Millipede
Chicobolus spinigerus
Paeromopus angusticeps
Paeromopus angusticeps
Texas Brown Tarantula
Aphonopelma hentzi
Portuguese Millipede
Ommatoiulus moreleti
Rusty Millipede
Trigoniulus corallinus
Desert Blonde Tarantula
Aphonopelma chalcodes
Desert Tarantula
Aphonopelma iodius
Striped Millipede
Ommatoiulus sabulosus
Narceus annularis
Narceus annularis
Desert Millipede
Orthoporus ornatus
Características
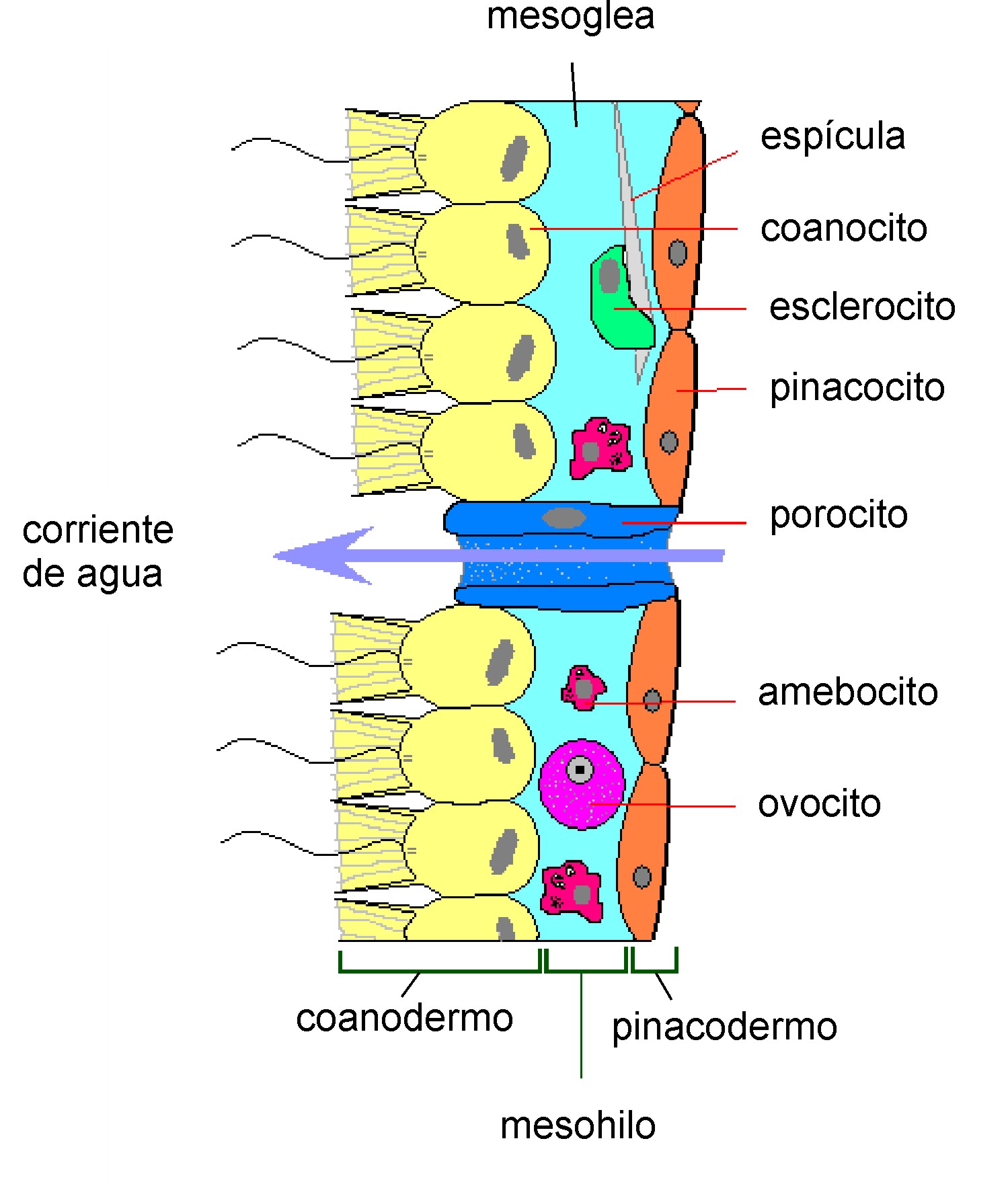
La movilidad es la característica más llamativa de los organismos de este reino, pero no es exclusiva del grupo, lo que da lugar a que sean designados a menudo como animales ciertos organismos, los llamados protozoos, que pertenecen al reino Protista. En el siguiente esquema se muestran las características comunes a todos los animales: Con pocas excepciones, la más notable la de las esponjas (filo Porifera), los animales presentan tejidos diferenciados y especializados. Estos incluyen músculos, que pueden contraerse para controlar el movimiento, y un sistema nervioso, que envía y procesa señales. Suele haber también una cámara digestiva interna, con una o dos aberturas. Los animales con este tipo de organización son conocidos como eumetazoos, en contraposición a los parazoos y mesozoos, que son niveles de organización más simples ya que carecen de algunas de las características mencionadas. Todos los animales tienen células eucariontes, rodeadas de una matriz extracelular característica compuesta de colágeno y glucoproteínas elásticas. Esta puede calcificarse para formar estructuras como conchas, huesos y espículas. Durante el desarrollo del animal se crea un armazón relativamente flexible por el que las células se pueden mover y reorganizarse, haciendo posibles estructuras más complejas. Esto contrasta con otros organismos pluricelulares como las plantas y los hongos, que desarrollan un crecimiento progresivo. - Organización celular: Eucariota y pluricelular. - Nutrición: Heterótrofa por ingestión (a nivel celular, por fagocitosis y pinocitosis), a diferencia de los hongos, también heterótrofos, pero que absorben los nutrientes tras digerirlos externamente. - Metabolismo: Aerobio (consumen obligatoriamente oxígeno). Sin embargo, recientemente se han descubierto varias especies del filo de animales marinos Loricifera, que tienen la particularidad de ser los primeros metazoos que hasta el momento se haya demostrado que vivan en un ambiente permanente de anaerobiosis, ya que no contienen mitocondrias, sino otros orgánulos. - Reproducción: Todas las especies animales se reproducen sexualmente (algunas solo por partenogénesis), con gametos de tamaño muy diferente (oogamia) y cigotos (ciclo diplonte). Algunas pueden, además, multiplicarse asexualmente. Son típicamente diploides. - Desarrollo: Mediante embrión y hojas embrionarias. El cigoto se divide repetidamente por mitosis hasta originar una blástula. - Estructura y funciones: Poseen colágeno como proteína estructural. Tejidos celulares muy diferenciados. Sin pared celular. Algunos con quitina. Fagocitosis, en formas basales. Ingestión con fagocitosis ulterior o absorción en formas derivadas ("más evolucionadas"), con capacidad de movimiento, etc. - Simetría: Excepto las esponjas, los demás animales presentan una disposición regular de las estructuras del cuerpo a lo largo de uno o más ejes corporales. Los tipos principales de simetría son la radial y la bilateral.
Funciones esenciales
Los animales llevan a cabo las siguientes funciones esenciales: alimentación, respiración, circulación, excreción, respuesta, movimiento y reproducción:
Alimentación
La mayoría de los animales no pueden absorber comida; la ingieren. Los animales han evolucionado de diversas formas para alimentarse. Los herbívoros comen plantas, los carnívoros comen otros animales; y los omnívoros se alimentan tanto de plantas como de animales. Los detritívoros comen material vegetal y animal en descomposición. Los comedores por filtración son animales acuáticos que cuelan minúsculos organismos que flotan en el agua. Los animales también forman relaciones simbióticas, en las que dos especies viven en estrecha asociación mutua. Por ejemplo, un parásito es un tipo de simbionte que vive dentro o sobre otro organismo, el huésped. El parásito se alimenta del huésped y lo daña.
Respiración
No importa si viven en el agua o en la tierra, todos los animales respiran; esto significa que pueden tomar oxígeno y despedir dióxido de carbono. Gracias a sus cuerpos muy simples y de delgadas paredes, algunos animales utilizan la difusión de estas sustancias a través de la piel. Sin embargo, la mayoría de los animales han evolucionado complejos tejidos y sistemas orgánicos para la respiración.
Circulación
Muchos animales acuáticos pequeños, como algunos gusanos, utilizan solo la difusión para transportar oxígeno y moléculas de nutrientes a todas sus células, y recoger de ellas los productos de desecho. La difusión basta porque estos animales apenas tienen un espesor de unas cuantas células. Sin embargo, los animales más grandes poseen algún tipo de sistema circulatorio para desplazar sustancias por el interior de sus cuerpos.
Excreción
Un producto de desecho primario de las células es el amoníaco, sustancia venenosa que contiene nitrógeno. La acumulación de amoniaco y otros productos de desecho podrían matar a un animal. La mayoría de los animales poseen un sistema excretor que bien elimina amoniaco o bien lo transforma en una sustancia menos tóxica que se elimina del cuerpo. Gracias a que eliminan los desechos metabólicos, los sistemas excretores ayudan a mantener la homeostasis. Los sistemas excretores varían, desde células que bombean agua fuera del cuerpo hasta órganos complejos como riñones.
Respuesta
Los animales usan células especializadas, llamadas células nerviosas, para responder a los sucesos de su medio ambiente. En la mayoría de los animales, las células nerviosas están conectadas entre sí para formar un sistema nervioso. Algunas células llamadas receptores, responden a sonidos, luz y otros estímulos externos. Otras células nerviosas procesan información y determinan la respuesta del animal. La organización de las células nerviosas dentro del cuerpo cambia dramáticamente de un fílum a otro.
Movimiento
Algunos animales adultos permanecen fijos en un sitio. Aunque muchos tienen movilidad. Sin embargo, tanto los fijos como los más veloces normalmente poseen músculos o tejidos musculares que se acortan para generar fuerza. La contracción muscular permite que los animales movibles se desplacen, a menudo en combinación con una estructura llamada esqueleto. Los músculos también ayudan a los animales, aun los más sedentarios, a comer y bombear agua y otros líquidos fuera del cuerpo.
Reproducción
La mayoría de los animales se reproducen sexualmente mediante la producción de gametos haploides. La reproducción sexual ayuda a crear y mantener la diversidad genética de una población. Por consiguiente, ayuda a mejorar la capacidad de una especie para evolucionar con los cambios del medio ambiente. Muchos invertebrados también pueden reproducirse asexualmente. La reproducción asexual da origen a descendiente genéticamente idénticos a los progenitores. Esta forma de reproducción permite que los animales aumenten rápidamente en cantidad.
Historia
Los animales han sido estudiados desde la antigüedad, y aún hoy, la clasificación animal se muestra cambiante, pues depende de los estudios que revelan constantemente información novedosa. Los grupos animales se definieron sobre la base de sus caracteres biológicos, morfológicos y ultraestructurales; sin embargo, la filogenia del siglo XXI está basada principalmente en el estudio filogenómico molecular del ADN mitocondrial, ribosómico y nuclear, lo que ha determinado también cambios importantes. La siguiente tabla, resume históricamente los sistemas de clasificación más notables, dando relevancia al descubrimiento de los principales supergrupos: La clasificación de Hyman (1940), que ha estado en vigencia hasta hace poco (Margulis & Chapman, 2009), ha sido invalidada por los estudios filogenéticos moleculares contemporáneos, ya que se demostró que grupos como los acelomados, pseudocelomados, celomados y esquizocelomados son en realidad grupos artificiales (polifiléticos).
Filos del reino animal
El reino animal se subdivide en una serie de grandes grupos denominados filos (el equivalente a las divisiones del reino vegetal); cada uno responde a un tipo de organización bien definido, aunque hay algunos de afiliación controvertida. En el siguiente cuadro, se enumeran los filos animales y sus principales características: En esta tabla no figuran los filos Echiura, Pogonophora, Sipuncula y Orthonectida los cuales han sido reclasificados en Annelida, y el filo Acanthocephala el cual fue reclasificado en Rotifera. El filo Myxozoa ha sido reclasificado en Cnidaria y el filo Monoblastozoa es de dudosa existencia.
Origen y documentación fósil
Mientras que en las plantas se conocen varias series de formas que conducen de la organización unicelular a la pluricelular, en el Reino Animal se sabe muy poco sobre la transición entre protozoos y metazoos. Dicha transición no está documentada por fósiles y las formas recientes supuestamente intermedias tampoco nos ayudan demasiado. En este campo de la transición pueden mencionarse, por una parte, a Proterospongia, coanoflagelado marino y planctónico que forma una masa gelatinosa con coanocitos en la parte exterior y células ameboides en el interior, y por otra al pequeño organismo marino Trichoplax adhaerens (filo placozoos) que forma una placa cerrada por epitelio pavimentoso en la parte dorsal y cilíndrico en la parte central, y presenta en la cavidad interior células en forma de estrella; se reproduce por yemas flageladas y huevos. Otra forma sencilla de metazoo es Xenoturbella, que vive sobre los fondos fangosos del mar. Tienen algunos centímetros de largo y forma de hoja, una boca ventral que conduce a un estómago en forma de saco. Entre la epidermis y el intestino existe una capa de tejido conjuntivo con un tubo muscular longitudinal y células musculares en el mesénquima; en la parte basal de la epidermis existe un plexo nervioso y en la parte anterior presenta un estatocisto; produce óvulos y espermatozoides, estos idénticos a los de diferentes metazoos primitivos. Su posición sistemática es incierta, habiéndose propuesto como miembro de un filo independiente (xenoturbélidos), a emplazar tal vez en la base de los deuteróstomos. Por lo que respecta a los mesozoos, ya no son considerados un estado de transición entre protistas y metazoos; su modo de vida parásito parece que les condujo a una reducción y simplificación extremas a partir de vermes acelomados. Por tanto, se debe recurrir a la morfología, fisiología y ontogenia comparadas de los metazoos para poder reconstruir esta etapa de la evolución. Los datos obtenidos con microscopía electrónica y análisis moleculares han apagado antiguas controversias sobre el origen de los metazoos. En este sentido, parece definitivamente rechazada la hipótesis sobre un origen polifilético; incluso los placozoos y los mesozoos, considerados a veces como originados directa e independientemente de los protistas, parecen a la luz de los nuevos datos claramente metazoos. Tres fueron las teorías sobre el origen de los metazoos: - Teoría colonial: Es la teoría universalmente aceptada que postula que los metazoos tuvieron un origen colonial a partir de los coanoflagelados, un pequeño grupo de Mastigóforos monoflagelados; algunos son individuales y otros coloniales. Dicha teoría se ve avalada tanto por datos moleculares (ARN ribosómico) como morfológicos (las mitocondrias y las raíces flagelares son muy semejantes en los metazoos y en los coanoflagelados, un cierto número de metazoos presenta células tipo coanocito, y los espermatozoides son uniflagelados en la mayor parte de ellos). Los seguidores de esta teoría incluyen al filo Choanoflagellatea en el reino animal, en contraposición al resto de animales, los metazoos. El antecesor de los metazoos, sería una colonia hueca y esférica de dichos flagelados; las células sería uniflageladas en su superficie externa; la colonia poseería un eje anteroposterior, nadando con el polo anterior hacia delante; entre las células somáticas existirían algunas células reproductoras. Este estado hipotético se ha denominado blastaea, y se cree que es el reflejo del estado de blástula que se produce en el desarrollo de todos los animales. Por tanto, esta teoría considera que los animales han evolucionado de protozoos flagelados. Sus parientes vivos más cercanos son los coanoflagelados, flagelados con la misma estructura que cierto tipo de células de las esponjas. Los análisis moleculares los sitúan en el supergrupo de los opistocontos, que también incluye a los hongos y a pequeños protistas parasitarios emparentados con estos últimos. El nombre viene de la localización trasera del flagelo en las células móviles, como en muchos espermatozoides animales, mientras que otros eucariontes tienen flagelos delanteros (acrocontos). El origen animal a partir de protozoos uniflagelados, así como su relación con los hongos, se puede graficar mediante las siguientes relaciones filogenéticasː - Teoría simbiótica: Una segunda hipótesis contemplaba la posibilidad que diferentes Protistas se hubiesen asociado simbióticamente originando un organismo pluricelular. Este es el origen que se presupone para las células eucariotas a partir de células procariotas. No obstante, no hay pruebas que respalden el origen simbiótico de los metazoos. - Teoría de la celularización: Otra teoría, que provocó profundas divergencias entre los zoólogos, es la que contemplaba a los turbelarios como los metazoos más primitivos y, por tanto, cuestiona el carácter ancestral de cnidarios y esponjas. Según esta hipótesis, los turbelarios derivarían de protistas ciliados multinucleados, por medio de celularización de los núcleos, lo que concuerda con el concepto de protozoo como organismo acelular. No obstante, hay muchos aspectos en contra de esta teoría, ya que no tiene en cuenta los criterios fundamentados en la embriología y da mucha más importancia a la organización del adulto.
Evolución del reino Animalia
Los primeros fósiles que podrían representar a animales corresponden a Otavia hallado en Namibia de entre 760-550 millones de años. Estos fósiles se interpretan como esponjas tempranas. Sin embargo estudios que usan relojes moleculares estiman el origen de los animales entre unos 850 millones de años durante la glaciación del Criogénico-Tónico. Los animales más antiguos que se conocen aparecen hacia el final del Precámbrico, hace alrededor de 580 millones de años, y se les conoce como vendobiontes o la biota del periodo Ediacárico. No obstante, son muy difíciles de relacionar con los fósiles posteriores. Algunos de estos organismos podrían ser los precursores de los filos modernos, pero también podrían ser grupos separados, y es posible que no fueran realmente animales en sentido estricto. Entre los primeros animales conocidos estarían Cyclomedusa, Charnia, Charniodiscus, Parvancorina, Annulatubus, Spriggina, etc. Aparte de ellos, muchos filos conocidos de animales hicieron una aparición más o menos simultánea durante el período Cámbrico, hace cerca de 570 millones de años. Todavía se discute si este evento, llamado explosión cámbrica, representa una rápida divergencia entre diferentes grupos o un cambio de condiciones que facilitó la fosilización. Algunos ejemplos serían Wiwaxia, Pikaia, Hallucigenia, Opabinia, etc. Entre los ancestros de grupos posteriores destaca Anomalocaris, del Cámbrico, como posible ancestro de diversos grupos de artrópodos, por su cuerpo segmentado, evolucionado de Opabinia y otros similares. Los cordados podrían tener relación con Pikaia. En cuanto a la evolución de los filos, tradicionalmente los animales se clasificaron por simetría y su nivel de complejidad en grupos como Radiata, Mesozoa, Acoelomata, Coelomata, y Pseudocoelomata que resultaron ser polifiléticos. Actualmente no está bien claro como fue el último ancestro común de todos los animales ("Urmetazoa"), todos los análisis moleculares respaldan la teoría colonial que afirma que los animales surgieron de la unión colonial de protozoos similares a los coanoflagelados que es la teoría ampliamente aceptada para explicar el origen de los animales. Parece poco probable que los animales hayan surgido de un único ancestro en común dado a la falta de homología entre los poríferos y los animales verdaderos (Eumetazoa), por lo que es probable que hayan surgido dos veces de las colonias de coanoflagelados. Los poríferos son muy diferentes de los eumetazoos puesto que carecen de tejidos, sistema nervioso, notocordio y son generalmente inmóviles, de hecho Haeckel las clasificaba en el reino Protista. Según los análisis moleculares el grupo más basal de los eumetazoos es Ctenophora lo que sugiere que los bilaterios, cnidarios y placozoos evolucionaron de ancestros similares a los ctenóforos. El origen y el ancestro de los bilaterios es un tema oscuro, pero los análisis moleculares han favorecido dos hipótesis clásicas: la hipótesis "Planulozoa" considera que los bilaterios surgieron de la derivación pedomórfica de la larva de un cnidario y según este punto de vista los gusanos acelomados (platelmintos, nemertinos y xenacelomorfos) serían los bilaterales más primitivos derivándose posteriormente de ellos celomados y pseudocelomados. Esta hipótesis fue respaldada por algunos estudios, pero a la vez criticada por otros, dado que los xenacelomorfos como platelmintos, son linajes muy reducidos, ha habido cambios constantes en el genoma y mutan más rápidamente que cualquier tipo de animal conocido, así que sugerir que estos linajes ocupan posiciones en los árboles filogenéticos podría significar caer en la atracción de ramas largas. Por el contrario, la otra hipótesis "Urbilateria" considera que el ancestro de los bilaterios fue un celomado (es decir, complejo) y con características de deuteróstomo derivándose los restantes celomados, los pseudocelomados y acelomados. Esta hipótesis es la más probable para el primer ancestro bilateral. Dentro de Bilateria dos clados han sido bien establecidos Protostomia respaldado por todos los análisis moleculares y Deuterostomia respaldado por la mayoría de los análisis moleculares, aunque algunos más recientes han cuestionado la monofilia de los deuteróstomos sugiriendo que son parafiléticos. Estos se definen por el desarrollo embrionario, en protóstomos la boca se desarrolla primero y el ano después, en cambio en los deuteróstomos es lo contrario el ano primero y la boca por último. Los análisis moleculares han demostrado que los pseudocelomados y acelomados (exceptuando los xenacelomorfos) son protóstomos reducidos y que evolucionaron de ancestros celomados en diferentes líneas evolutivas por lo que son grupos polifiléticos. Estos filos se han clasificados junto con los anélidos, moluscos, artrópodos, en los clados Spiralia y Ecdysozoa respectivamente. Antiguamente se creía que los anélidos estaban más estrechamente con los artrópodos y que los lofoforados con quetognatos eran deuteróstomos, pero los análisis moleculares han demostrado que estos últimos pertenecen Spiralia junto con anélidos, moluscos, etc. La posición de los entoproctos que fue incierta durante muchos también se resolvió en Spiralia. El clado Spiralia contiene los (moluscos, anélidos, lofoforados, quetognatos, platelmintos, nemertinos, gastrotricos, rotíferos, entoproctos, ciclióforos, gnatostomúlidos, micrognatozoos, rombozoos), y el clado Ecdysozoa (artrópodos, tardígrados, onicóforos, nematodos, nematomorfos, quinorrincos, priapúlidos y loricíferos). Estos clados se han definido por la segmentación espiral del huevo y la muda de cutícula (ecdisis). En los análisis moleculares los filos pseudocelomados ocupan las posiciones más basales en Spiralia y Ecdysozoa lo que sugiere que los ancestros de los celomados protóstomos (ej.- artrópodos, moluscos, anélidos, etc.) fueron pseudocelomados. El ancestro de los platelmintos y nemertinos reducidos pudo haber sido un celomado emparentado con los anélidos. Por otra parte el supergrupo Deuterostomia incluye a los (cordados, equinodermos, hemicordados y xenacelomorfos), de estos filos los equinodermos y hemicordados forman un clado bien soportado por los análisis moleculares y morfológicos, el cual puede estar relacionado con los xenacelomorfos, a su vez los cordados conforman la rama más basal. En la evolución de los animales ha habido un grupo de animales que han sido difíciles de relacionar y que tradicionalmente se consideraron los más primitivos por su carácter simple se trata de los mesozoos (placozoos, ortonéctidos y rombozoos), sin embargo se ha demostrado que son formas totalmente reducidas por el parasitismo y que de hecho no tienen ningún parentesco. Los placozoos serían radiados reducidos, los ortonéctidos anélidos reducidos y los rombozoos descendientes de un linaje espiralio (Spiralia) reducido probablemente emparentado con los anélidos, platelmintos y nemertinos. Algunos filos fueron clasificados en otros filos como Echiura, Pogonophora, Sipuncula y Orthonectida que si bien no tienen la segmentación y metamerización típica de los anélidos la perdieron durante su evolución. Acanthocephala tradicionalmente considerado un filo separado se clasifican en los rotíferos, ya que evolucionaron de rotíferos según los análisis moleculares y morfológicos. También algunos animales llegaron al punto de ser unicelulares como Myxozoa que anteriormente se clasificaba como protistas. Los análisis moleculares han demostrado que Myxozoa evolucionó de un ancestro cnidario pluricelular emparentado con Polypodiozoa.

Filogenia clásica
El siguiente cladograma representa las relaciones filogenéticas entre los diversos filos de animales. Está basada en la segunda edición de Brusca & Brusca (2005); se trata de una hipótesis filogenética "clásica" en la que se reconocen los grandes clados admitidos tradicionalmente (pseudocelomados, articulados, etc.) y asume la teoría colonial como la explicación sobre el origen de los metazoos. Investigaciones más recientes ofrecen una visión algo diferente. Según el punto de vista que se acaba de exponer, los bilaterales se subdividen en cuatro grandes linajes: Las modernas técnicas de secuenciación de bases del ADN, ARN y proteínas junto con la metodología de la cladística han permitido reinterpretar las relaciones filogenéticas de los distintos filos animales, lo que ha conducido a una revolución en la clasificación de los mismos. Actualmente la mayoría de los zoólogos aceptan las nuevas clasificaciones. Los animales bilaterales parecen pertenecer a uno de estos tres linajes: - Protóstomos acelomados - Protóstomos esquizocelomados - Protóstomos pseudocelomados - Deuteróstomos - Deuteróstomos - Protóstomos - Ecdisozoos - Espiralios
Filogenia actual
La filogenia de los animales al igual que otras filogenias es un tema no resuelto y todavía no se ha alcanzado un consenso amplio en cuanto a la relación filogenética de los determinados filos debido a la gran cantidad de hipótesis alternativas que se sugieren en los diversos artículos científicos. La filogenética molecular ha producido una revolución en las relaciones filogenéticas de los animales reemplazando muchas hipótesis tradicionales. Tampoco existe muchos datos morfológicos, fósiles y biogeográficos fiables como para tomar hipótesis alternativas. Especialmente las relaciones filogenéticas de los superfilos Spiralia y Ecdysozoa son tan ambiguas que no se puede llegar a un consenso sobre cual es la filogenia correcta. En cuanto a grupos extintos Vendobionta es un completo enigma evolutivo, tentativamente se le puede considerar en la base de la filogenia animal, aunque solo comparte con las esponjas su simplicidad y hábitat bentónico, además las esponjas serían menos antiguas que aquel. Recientemente se ha sugerido que los filos Rotifera, Rhombozoa, Ctenophora, Placozoa, Entoprocta, Platyhelminthes, Gastrotricha, Xenacoelomorpha, Chaetognatha y Gnathostomulida tienen tasas de evolución genética, lo que impide reconstruir el árbol filogenético y llevan a las atracciones de ramas largas. Análisis moleculares recientes (2019) utilizando especies de evolución lenta y otros estudios que intentan evitar el error sistemático han dado resultados posiblemente correctos. A diferencia de otros análisis moleculares convencionales que llevan a resultados contradictorios, estos análisis están de acuerdo con los análisis morfológicos y paleontológicos. A continuación se grafica el posible árbol de diversificación animal según sus conclusiones:
Un árbol filogenético alternativo
A continuación se muestra un árbol filogenético alternativo publicado en el libro "The Invertebrate Tree Of Life" (2020) es el propuesto por Gonzalo Giribet. Los nodos marcados son defendidos por el autor:
Bibliografía
- Conway, Morris, S. 1993. The fossil record and the early evolution of the Metazoa. Nature 361:219-225. An important summary correlating fossil and molecular evidence. - Hickman, C. P., Ober, W. C. & Garrison, C. W. 2006. Principios integrales de zoología, 13.ª edición. McGraw-Hill-Interamericana, Madrid (etc.), XVIII+1022 pp. ISBN 84-481-4528-3. - Storer, Tracy. General Zoology. 6th edition. MC. Graw Hill Book Company, Inc.